
Turfgrass chemistry and annual bluegrass weevil survival
The annual bluegrass weevil (Listronotus maculicollis) remains one of the most destructive and costly insect pests affecting closely mown cool-season turf across eastern North America. Historically confined to the northeastern United States and strongly associated with annual bluegrass (Poa annua), the pest expanded rapidly beginning in the late 1990s.
Today, damaging populations extend from Canadian provinces to the Mid-Atlantic and Midwest, with recent reports from Arkansas, Kentucky, Wisconsin, Indiana, Michigan, Nebraska, Georgia, Kansas and Illinois. Its spread into regions dominated by warm-season turf raises new questions about host suitability and future risk.
Annual bluegrass remains the preferred and most susceptible host. Perennial ryegrass can support populations but rarely sustains severe injury, while creeping bentgrass functions as a secondary host where larvae develop more slowly and with reduced survival. This reduced fitness may be linked to plant defense chemistry.
Creeping bentgrass contains higher levels of jasmonic acid, a defense hormone that stimulates production of anti-herbivory compounds such as phenols and flavonoids. These compounds reduce plant nutritional quality, disrupt insect digestion and development and may improve turf tolerance to feeding injury. Similar flavonoid-rich defenses in zoysiagrass and bermudagrass are known to suppress fall armyworm development, suggesting comparable mechanisms could limit weevil survival.
Interest in host plant resistance as a management strategy is growing. Unlike annual bluegrass, many turf species have been bred for stress tolerance, disease resistance and improved performance, though rarely for insect resistance. Darker green cultivars often contain higher levels of protective compounds, suggesting that pigment intensity may serve as a biomarker of insect tolerance. Conversely, decades of breeding bentgrass for aesthetics may have reduced defensive traits.
Ongoing research is evaluating cool- and warm-season turfgrasses to determine their suitability for larval development and to identify nutritional traits linked to resistance. These findings could guide breeding programs and support nonchemical strategies for managing annual bluegrass weevil in an expanding geographic range.
Materials and methods
Larval recovery and survivorship on cool- and warm-season turfgrasses: Adult annual bluegrass weevils were collected from golf course fairways, collars and tees in central Pennsylvania in June 2022 and 2023 using a reverse leaf-blower vacuum. Adults were transported to Penn State’s Turfgrass Entomology Laboratory, sorted by sex and viability and held in climate-controlled chambers (14 h light at 70 degrees F; 10 h dark at 57 degrees F) without supplemental food until use.
No-choice assays were conducted by introducing two males and two females into intact 4.25-by-4.0-inch turf cores (10.8 by 10.2 centimeters) of annual bluegrass, creeping bentgrass (“Penncross” and “Penn-A4”) perennial ryegrass, zoysiagrass (“Zeon” Zoysia matrella) and hybrid bermudagrass (“Latitude 36” Cynodon dactylon x C. transvaalensis).
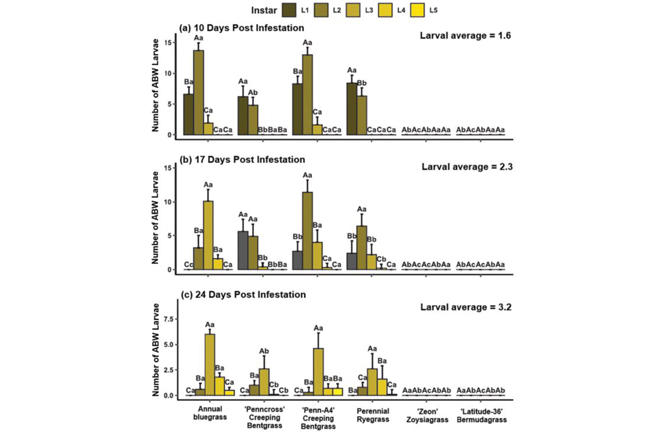
Adults were removed after seven days to prevent continued egg laying. Larvae were extracted 10, 17 and 24 days after infestation using heat-driven Berlese funnels. Recovered larvae were counted and staged by head capsule width to determine instar and an average larval instar value was calculated to compare development rates and survivorship among hosts.
Turfgrass maintenance and extract production: Turf cores were collected from research nursery plots protected from natural infestations and maintained without fertility or plant growth regulators. Cores were stored under controlled environmental conditions, watered as needed and trimmed to a uniform 0.8-inch (20-mm) height of cut before infestation. Clippings collected during the study were macerated and extracted in methanol to obtain soluble plant compounds and metabolites for chemical analysis.
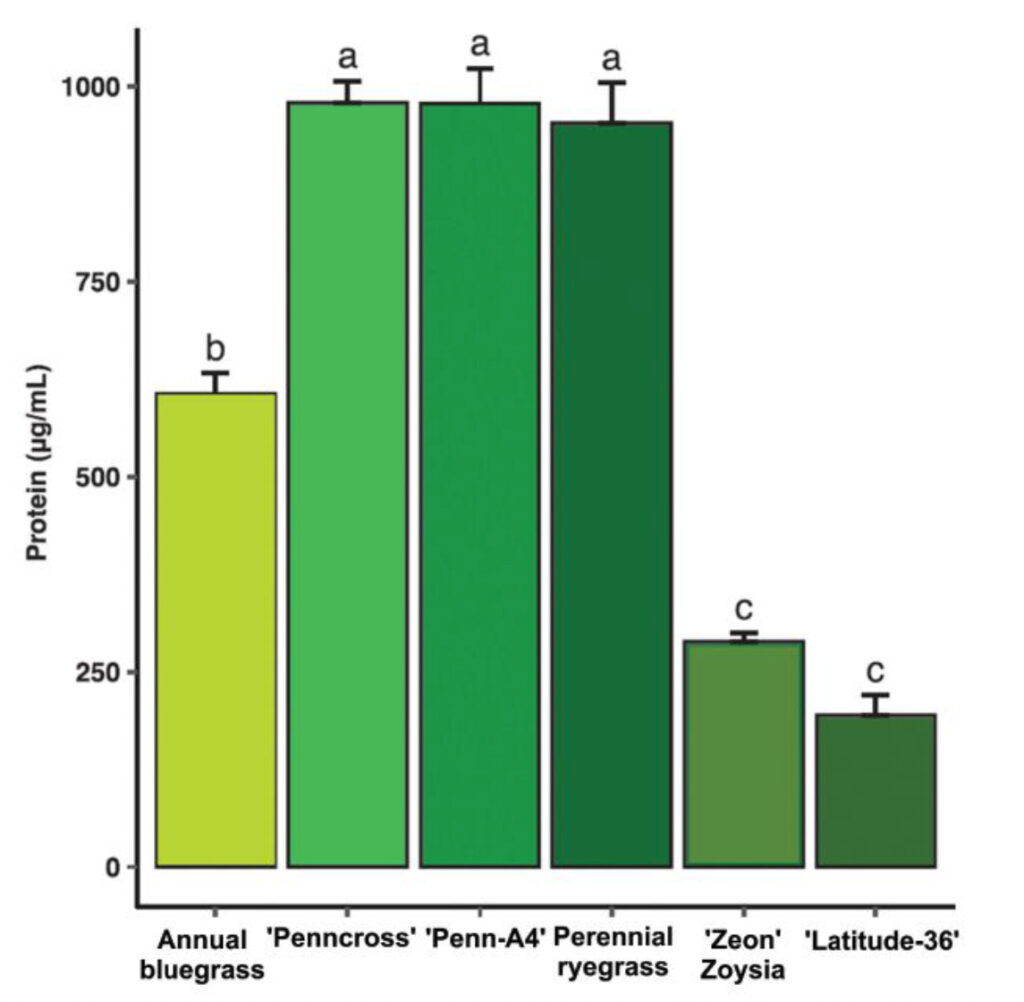
Soluble protein content: Soluble protein concentrations were quantified from turf extracts using a microplate spectrophotometer assay. Diluted extracts were analyzed in replicated trials to determine protein availability, a factor influencing insect growth and host suitability.
![(a) Total carotenoid (mg/100 mL fresh weight [FW]) and (b) chlorophyll (mg/100 mL FW) content of annual bluegrass, bentgrass, perennial ryegrass, zoysiagrass and hybrid bermudagrass. Columns with the same lowercase letter denote no significant differences in carotenoid (a) and chlorophyll (b) content between turfgrass species/cultivars at α =0.05.](https://www.golfdom.com/wp-content/uploads/2026/04/figure3-632x1024.jpg)
Total carotenoid and chlorophyll: Chlorophyll and carotenoid pigments were extracted and quantified spectrophotometrically to assess plant physiological status and potential links to defensive chemistry. Absorbance readings were used to calculate pigment concentrations.
Total phenol and flavonoid: Total phenols and flavonoids, key anti-herbivory compounds, were measured using standard colorimetric assays. Phenols were expressed as gallic acid equivalents (GAE/L), and flavonoids as quercetin equivalents (QE/g dry weight [DW]). These compounds can reduce plant palatability, interfere with digestion and impair insect development.
Statistical analysis: Analysis of variance (ANOVA) was used to evaluate turfgrass effects on larval survivorship and development and to compare nutritional and defensive compounds among species and cultivars. When significant differences were detected, means were separated using Tukey’s test at α = 0.05.
Results
Larval recovery and survivorship: Larval recovery was significantly greater from annual bluegrass and Penn-A4 at 10 (Figure 1a) and 17 (Figure 1b) days post infestation (DPI). At 24 DPI, perennial ryegrass hosted similar densities of larvae as annual bluegrass and Penn-A4, which were significantly greater than Penncross and both warm-season grasses (Figure 1c).
Larvae reared on Penncross experienced significantly longer development time at 10, 17 and 24 DPI compared to that of annual bluegrass. No larvae were recovered from either Latitude 36 bermudagrass or Zeon zoysiagrass in either year of the study.
Soluble protein content: Both creeping bentgrass cultivars and perennial ryegrass contained significantly greater protein content than annual bluegrass and warm-season grasses (Figure 2). Annual bluegrass soluble protein concentrations were significantly higher than those of both warm-season grasses. However, protein content in annual bluegrass was significantly lower than that of the cool-season turfgrasses.
Total carotenoid and chlorophyll: Penncross and perennial ryegrass contained significantly greater carotenoid (mg/100 mL fresh weight [FW]) content than annual bluegrass, Penn-A4, Latitude 36 bermudagrass or Zeon zoysiagrass (Figure 3a). Poa annua carotenoid content was significantly lower among cool-season grasses but significantly higher than Latitude 36 bermudagrass.
Of the warm-season turf assessed, Zeon zoysiagrass carotenoid concentrations were significantly more elevated than Latitude 36 bermudagrass. Both creeping bentgrasses, perennial ryegrass and Zeon zoysiagrass possessed significantly higher chlorophyll levels than those of annual bluegrass and Latitude 36 bermudagrass (Figure 3b).
![(a) Total phenol (mg gallic acid equivalents [GAE]/L) and flavonoid (mg quercetin equivalent [QE]/g DW) content of annual bluegrass, bentgrass, perennial ryegrass, zoysiagrass and hybrid bermudagrass. Columns with the same lowercase letter denote no significant differences in phenol (a) and flavonoid (b) content between turfgrass species/cultivars at α =0.05.](https://www.golfdom.com/wp-content/uploads/2026/04/figure4-600x1024.jpg)
Total phenol and flavonoid: Penncross bentgrass and perennial ryegrass total phenol (mg GAE/L) levels were significantly higher than those found in annual bluegrass, Latitude 36 bermudagrass or Zeon zoysiagrass. Notably, annual bluegrass phenol levels were not statistically different from those of both warm-season turfgrasses evaluated (Figure 4a).
Penncross bentgrass and perennial ryegrass had significantly greater total flavonoid content than annual bluegrass, Penn-A4 bentgrass and Zeon zoysiagrass. Annual bluegrass demonstrated the lowest flavonoid content, while Penncross bentgrass exhibited the highest total flavonoid concentrations (Figure 4b).
Discussion
This study provides the first comprehensive evaluation of turfgrass nutritional chemistry and host suitability for annual bluegrass weevil larval development. It offers new insights into resistance mechanisms across cool- and warm-season turf species. A key finding was the failure to recover first-instar larvae from Zeon zoysiagrass or Latitude 36 hybrid bermudagrass under no-choice conditions, suggesting these warm-season grasses may not support oviposition or larval development.
Whether this resistance is structural, physiological or both remains unclear. Warm-season grasses are known to contain silica in leaf tissues that increase rigidity and deter feeding, and elevated fiber, lignin and silicon levels can reduce digestibility and insect performance.
However, unlike tolerant cool-season hosts, phenols and flavonoids were relatively low in these warm-season grasses, indicating that chemical defenses common in cool-season grass tolerance may not explain resistance in warm-season turf. Further research into chemical and structural defenses could clarify tolerance mechanisms in warm-season species.
Among cool-season turfgrasses, larval survivorship and development differed significantly. Creeping bentgrass Penncross demonstrated consistent tolerance, supporting fewer larvae and reduced survivorship compared with the newer cultivar Penn-A4. This supports previous observations that modern bentgrass breeding focused on aesthetics, density and disease resistance may have inadvertently reduced insect tolerance, increasing susceptibility to levels approaching those of annual bluegrass.
Plant nutritional composition appears closely linked to host suitability. Both bentgrass cultivars and perennial ryegrass contained greater soluble protein concentrations than annual bluegrass and warm-season grasses. Protein availability influences insect growth and host specialization, but defensive compounds, such as phenols, including flavonoids, can bind proteins and reduce digestibility, thereby limiting the nutritional value for feeding larvae.
Larvae reared on Penncross experienced the greatest decline in survivorship among all turfgrasses tested. This cultivar contained higher levels of chlorophyll, carotenoids, phenols, flavonoids and protein than annual bluegrass, the most susceptible host.
Phenols and flavonoids can deter feeding, disrupt digestion, reduce nutrient availability and interfere with insect development and hormone regulation. These compounds also contribute to darker pigmentation, and the study confirmed a strong relationship between darker green color and elevated defensive chemistry. Pigments such as chlorophyll and carotenoids, often selected for turf aesthetics, may therefore serve as visual indicators of insect tolerance.
Annual bluegrass exhibited the lowest flavonoid concentrations and remains highly susceptible to weevil feeding. Its susceptibility may also be linked to defense signaling dominated by salicylic acid pathways, which are associated with pathogen defense. It can suppress herbivory-related defenses and reduce the production of insect-deterring compounds. In contrast, creeping bentgrass appears to rely more heavily on jasmonic acid-mediated herbivory defenses.
The findings reinforce earlier observations that Penncross consistently supports lower larval densities than newer bentgrass cultivars, suggesting insect tolerance traits may have been lost during selection for other desirable characteristics. Identifying defensive biomarkers, particularly phenols and flavonoids, and understanding the structural defenses of warm-season grasses could help breeders develop cultivars with improved insect resistance.
Overall, this research demonstrates that turfgrass susceptibility to annual bluegrass weevil is strongly influenced by plant chemistry, defense signaling and breeding history. Turfgrasses with elevated pigments and defensive compounds exhibit greater tolerance, while warm-season grasses may resist infestation through structural defenses. These insights provide a foundation for breeding insect-resistant cultivars and advancing nonchemical management strategies for this expanding pest.
This article was adapted from the following source: Simard, A., and McGraw, B. A. (2025). Characterization of cool- and warm-season turfgrass host suitability to annual bluegrass weevil (Listronotus maculicollis, Kirby). Crop Science, 65, e21363. https://doi.org/10.1002/csc2.21363
For additional information, contact Audrey Simard, Pennsylvania State University, at audreysimard@envu.com.
Subscribe to Golfdom

If you enjoyed this article, subscribe to Golfdom to receive more articles just like it.



